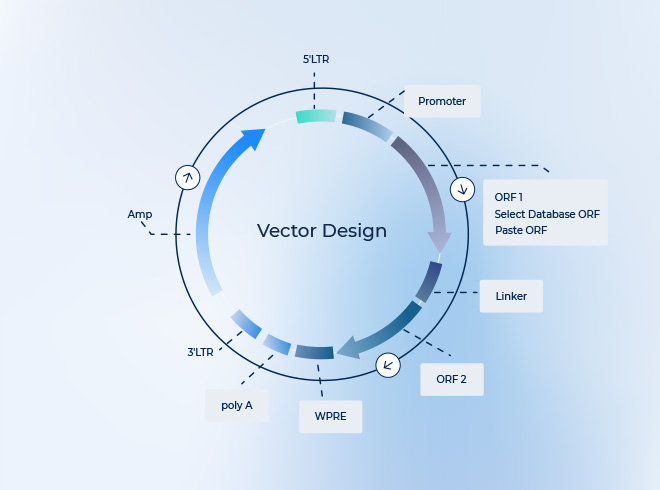
AAV(腺相关病毒,adeno-associated virus)质粒通常由以下几个关键结构元件组成:
1. 反向末端重复序列(ITRs – Inverted Terminal Repeats)
-
位置与功能:位于质粒的最两端,是AAV基因组的核心顺式作用元件。
-
必要性:是AAV基因组复制、从生产质粒上被“剪切”下来、以及最终被包装到病毒衣壳内所必需的唯一来自野生型AAV的序列。没有ITRs,就无法生产出rAAV。
-
结构:形成发夹结构的回文序列,长约145个碱基对。
2. 启动子(Promoter)
-
功能:控制下游目的基因的转录起始,决定基因在什么细胞、什么时候、以多强的水平进行表达。
-
类型:
-
通用型强启动子:如CMV(巨细胞病毒立即早期启动子)、CAG(复合启动子,包含CMV增强子和鸡β-肌动蛋白启动子),能在大多数细胞类型中驱动高强度表达。
-
组织/细胞特异性启动子:如hSyn(人突触蛋白启动子,用于神经元)、GFAP(胶质纤维酸性蛋白启动子,用于星形胶质细胞)、Alb(白蛋白启动子,用于肝细胞),可限制基因在特定细胞中表达。
-
3. 目的基因(Gene of Interest, GOI)
-
功能:这是整个载体的有效载荷(Payload),即你希望递送并表达的基因(如治疗性基因、报告基因如GFP、或shRNA等)。
4. 终止信号/多聚腺苷酸化信号(Polyadenylation Signal, PolyA)
-
功能:位于目的基因的下游,为mRNA转录提供终止信号,并在其3’末端添加一串多聚腺苷酸(polyA)尾。这对于mRNA从细胞核转运到细胞质、以及其稳定性至关重要。
-
常见类型:SV40 polyA、BGH(牛生长激素)polyA、合成polyA等。
5. 增强表达/调控的辅助元件(可选但常用)
为了优化表达,通常会在表达框架中加入一些辅助元件:
-
WPRE(土拨鼠肝炎病毒转录后调控元件):位于终止子之前,能显著增强mRNA的稳定性和核输出,从而提高蛋白表达水平。
-
内含子(Intron):如来自鸡β-肌动蛋白基因的内含子。内含子的存在可以大大增强pre-mRNA的剪切和成熟效率,从而提升基因表达水平。
-
miRNA靶位点(miRNA Target Sites):用于在特定细胞类型中“沉默”或“降低”转基因的表达。例如,在靶向神经元的载体中加入肝细胞特异性miRNA的靶位点,可以“脱靶”表达,使病毒在进入肝细胞时被沉默,增强特异性。
6. 质粒骨架(Bacterial Backbone)
这部分不会被包装到rAAV病毒颗粒中,其唯一作用是为了在实验室里用大肠杆菌大量扩增这个质粒。
-
细菌复制起点(ori):如pUC ori,允许质粒在细菌中进行高水平复制,从而获得高产量的质粒DNA。
-
抗生素抗性基因:如氨苄青霉素抗性基因(AmpR)或卡那霉素抗性基因(KanR),用于在细菌培养过程中筛选出成功转化了该质粒的菌落。
在生产rAAV病毒时,AAV质粒与另外两个提供反式作用蛋白的辅助质粒(包装质粒提供Rep/Cap蛋白,辅助质粒提供腺病毒基因如E2A/E4/VA)共转染到生产细胞(如HEK293细胞)中。细胞机器会识别载体质粒上的ITR,并将两个ITR之间的所有序列剪切、复制并包装到由Cap蛋白构成的病毒衣壳中,形成有感染能力的rAAV病毒颗粒。
关于派真
作为一家专注于AAV 技术十余年,深耕基因治疗领域的CRO&CDMO,派真生物可提供从载体设计、构建到 AAV、慢病毒和 mRNA 服务的一站式解决方案。凭借深厚的技术实力、卓越的运营管理和高标准的服务交付,我们为全球客户提供一站式CMC解决方案,包括从早期概念验证、成药性评估到IIT、IND及BLA的各个阶段。
凭借我们独立知识产权的π-alphaTM 293 细胞AAV高产技术平台,我们能将AAV产量提高多至10倍,每批次产量可达1×10¹⁷vg,以满足多样化的商业化和临床项目需求。此外,我们定制化的mRNA和脂质纳米颗粒(LNP)产品及服务覆盖药物和疫苗开发的各个阶段,从研发到符合GMP的生产,提供端到端的一站式解决方案。