AAV感染效率低是一个多因素导致的问题,主要原因可以分为以下几个方面:
1. 细胞自身因素
-
细胞表面受体表达水平:AAV进入细胞的第一步是结合细胞表面的初级受体(如AAV2需要硫酸肝素蛋白聚糖HSPG)和共受体(如αVβ5整合素、成纤维细胞生长因子受体FGFR1等)。如果目标细胞不表达或低表达这些特异性受体,AAV就无法有效吸附和进入细胞。这是决定细胞嗜性 的关键。
-
细胞内吞与胞内运输效率:AAV通过网格蛋白介导的内吞作用进入细胞后,需要经历从早期内体到晚期内体,再到核内体的复杂运输过程。这个过程中,大部分病毒颗粒会被运输到溶酶体并被降解,只有少部分能成功“逃逸”到细胞质中。
-
核膜屏障:AAV的基因组需要进入细胞核才能进行转录。AAV的衣壳大小约20-25nm,而核孔复合物的有效直径通常小于10nm。因此,完整的AAV病毒颗粒很难直接通过核孔。它需要依赖核膜上不稳定的“临时孔洞”(如在细胞分裂时或某些特定条件下)或者一个复杂且效率不高的核内体释放和入核机制。
2. 病毒载体因素
-
血清型选择不当:不同的AAV血清型(如AAV1, 2, 5, 6, 8, 9等)对不同的组织和细胞具有天然的嗜性差异。例如,AAV9易于穿过血脑屏障感染中枢神经系统,而AAV8对肝脏有高亲和力。如果选择的血清型与目标组织不匹配,感染效率会非常低。
-
衣壳免疫原性:虽然AAV的免疫原性远低于腺病毒,但人群中仍普遍存在针对不同AAV血清型的中和抗体。这些抗体可以识别并结合AAV衣壳,在病毒进入目标细胞前就将其清除,从而完全阻断感染。
-
病毒滴度不足或质量不佳:制备的病毒原液如果实际感染性滴度(IU/mL)低于理论值,或者在生产纯化过程中病毒衣壳受损,都会直接导致功能性病毒颗粒数量不足,感染效率下降。
3. 细胞内及机体内的限制性因素
-
细胞内降解:如上所述,大部分AAV在胞内运输过程中被困在核内体/溶酶体途径中被降解,这是限制感染效率的一个主要“瓶颈”。
-
基因组第二链合成(针对部分血清型):某些AAV血清型(如AAV2)的基因组是单链DNA。它进入细胞核后,需要依赖宿主细胞的DNA聚合酶合成第二条互补链,形成双链DNA模板,才能启动基因表达。这个过程效率较低且速度慢,是导致转基因表达延迟和效率降低的重要原因。这也是自我互补型AAV(scAAV)被开发出来的原因,它绕过了这一步骤,能更快、更强地表达基因。
-
机体免疫清除:在体内实验中,除了预存的中和抗体,先天免疫系统和细胞免疫系统也会被激活。巨噬细胞等会吞噬和清除血液循环中的AAV颗粒,而细胞免疫(如CD8+ T细胞)可能会攻击和清除那些成功表达了外源转基因的细胞。
总结与对策
为了提高AAV的感染效率,研究人员采取了多种策略:
| 问题根源 | 解决策略 |
| 受体限制 | • 选择合适血清型:根据目标组织选择嗜性最佳的血清型。 |
| • 衣壳工程化改造:通过定向进化或理性设计,改造衣壳蛋白,使其能识别新的受体,增强对特定细胞的靶向性。 | |
| 入核和基因组转换障碍 | • 使用自我互补型AAV(scAAV):绕过第二链合成,提高表达速度和效率。 |
| • 使用高感染复数(MOI):在体外实验中,通过增加病毒颗粒与细胞的数量比来强行提高感染率。 | |
| 细胞内降解 | • 添加辅助药物:在感染时加入组蛋白去乙酰化酶抑制剂(如VPA)或蛋白酶体抑制剂(如Bortezomib),可以抑制病毒基因组在核内的沉默或减少其在胞内的降解。 |
| 免疫清除 | • 血清型筛选:选择患者体内预存中和抗体率较低的血清型。 |
| • 空壳病毒去除:在生产过程中尽可能去除不含基因组的“空壳”病毒颗粒,因为它们会无效消耗抗体和受体。 | |
| • 使用免疫抑制剂:在临床治疗中短期使用免疫抑制剂来控制T细胞反应。 |
AAV感染效率低是一个系统性问题,需要从病毒载体设计(衣壳、基因组)、目标细胞特性、以及体内外环境等多个层面进行综合分析和优化。
关于派真
作为一家专注于AAV 技术十余年,深耕基因治疗领域的CRO&CDMO,派真生物可提供从载体设计、构建到 AAV、慢病毒和 mRNA 服务的一站式解决方案。凭借深厚的技术实力、卓越的运营管理和高标准的服务交付,我们为全球客户提供一站式CMC解决方案,包括从早期概念验证、成药性评估到IIT、IND及BLA的各个阶段。
凭借我们独立知识产权的π-alphaTM 293 细胞AAV高产技术平台,我们能将AAV产量提高多至10倍,每批次产量可达1×10¹⁷vg,以满足多样化的商业化和临床项目需求。此外,我们定制化的mRNA和脂质纳米颗粒(LNP)产品及服务覆盖药物和疫苗开发的各个阶段,从研发到符合GMP的生产,提供端到端的一站式解决方案。

相关服务
-
AAV病毒包装
- 6天极致交付
- 100+ 血清型,60+ 质量控制检测
- 已交付 50,000+ AAV,可放大至 GMP 级别
-
AAV病毒包装 – 科研级
- 6天极致交付
- 100+ 血清型,60+ 质量控制检测
- 已交付 50,000+ AAV,可放大至 GMP 级别
-
AAV病毒包装 – NHP级
- 严控内毒素,防污染管控
- ddPCR 精准定量,基因组完整性验证
- 卓越的一致性,安全性与有效性双重提升
-
AAV病毒包装 – HT级
- 最快 7 个工作日极速交付
- 采用 qPCR 滴度测定,具备高性价比
- 优化工艺以在细胞培养中实现卓越性能
-
AAV分析检测服务
- 全面的 AAV 检测:滴度、纯度、安全性等一应俱全
- 结果快速可靠,专家全程指导
-
AAV血清型筛选
- 定向进化 + 理性设计,精准打造理想衣壳
- 体内数据可信赖,候选分子轻松筛选
- 项目专属保密,突变体逐一验证
-
AAV文库服务
- 定制化 AAV 文库服务,提升基因递送的精确性
- 最大限度减少脱靶效应和先天性免疫反应
- 提高基因治疗研究的有效性
-
AAV现货
- 现货直发,到手即用
- 品类齐全,覆盖多元化科研需求
- 严苛质控,品质稳定可靠
-
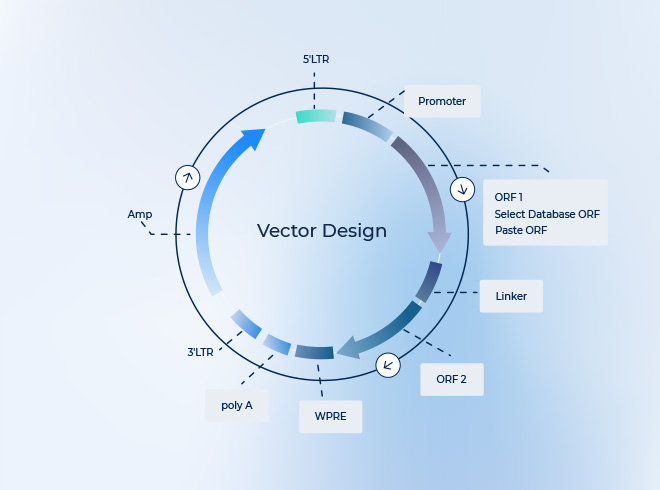
AAV载体设计与构建
- 定制化AAV质粒,实现高效基因递送
- 通过piVector轻松设计,集成特定应用所需元件
- 支持CRISPR、shRNA及其他类型的AAV载体