
基因承载着遗传信息,定义了生命的多样性和复杂性。基因编辑是理解和改造生命的关键技术,在生物学研究和生物产业发展中发挥着重要作用。历代基因编辑工具,如巨核酸酶、ZFN、TALEN均以蛋白质为基础,识别和切割DNA,编辑位点的重编程较为困难。目前被广泛使用的CRISPR-Cas工具,是RNA引导的蛋白核酸酶,通过引导RNA的间隔序列来识别DNA,具有很好的编辑位点重编程能力,但依然存在着PAM序列限制、分子量大、蛋白免疫原性等多种问题。
2024年2月1日,清华大学生命学院刘俊杰(Jun-Jie Gogo Liu)课题组在国际顶尖学术期刊 Science 发表了题为:Hydrolytic endonucleolytic ribozyme (HYER) is programmable for sequence-specific DNA cleavage 的研究论文【1】。
该研究报道了一种新型基因编辑工具——HYER(水解型内切核酶),仅由具有催化活性的RNA(核酶)构成,HYER可序列特异性地切割RNA和DNA底物,并在哺乳动物细胞基因组中产生位点特异性的编辑。研究团队还通过理性设计,获得了工程化HYER,具有更高的特异性和灵活性。这也是科学界首次报道具有DNA水解切割能力的核酶。
HYER对底物的识别和切割仅依赖于RNA分子,而无需蛋白核酸酶参与,由于具有良好的可编程性,HYER有望成为继CRISPR之后的新一代基因编辑底盘工具。


刘俊杰研究员
刘俊杰,2011本科毕业于四川大学,2017年博士毕业于清华大学,此后曾在诺奖得主、CRISPR基因编辑先驱 Jennifer Doudna 教授实验室进行博士后研究,在此期间作为第一作者发表 Nature 论文【2】,开发了一种基于CRISPR-Cas12e的小型化基因编辑工具——CasX,CasX转化的基因编辑疗法公司Scribe Therapeutics已完成超1亿美元融资,并与礼来、赛诺菲等制药巨头达成大额合作。
2020年10月,刘俊杰加入清华大学生命学院,成立了独立实验室。2023年1月,刘俊杰团队在 Cell Research 期刊发表论文【3】,开发了基于CRISPR-Cas12l的小型化基因编辑工具——Casπ。2023年6月,刘俊杰团队在Cell期刊发表论文【4】,解析了R2逆转座子在基因组上跳跃和切割DNA的分子机制,为开发基于逆转座子的新型基因编辑工具奠定了基础。
值得一提的是,在这篇Cell论文中,刘俊杰团队还将来自低等真核生物的R2逆转座子与其祖先(原核生物第二类内含子,GII intron)及哺乳动物中的LINE-1逆转座子进行了比较,发现第二类内含子向R2逆转座子及进一步向LINE-1逆转座子进化的过程中,RNA的结构性组分逐渐减少并被编码区域所取代,并且催化功能逐渐从以RNA为主导过渡到以蛋白质为主导。
1980年代,Thomas Cech 和 Sidney Altman 等人发现并证明了RNA也可以发挥催化作用,即所谓的核酶(Ribozymes),他们也因此获得了1989年的诺贝尔化学奖。
第二类内含子(GII intron)逆转座子,特别是它们编码的内含子RNA,是一类能够在宿主基因组通过“复制-粘贴”进行逆转座的可移动遗传元件,该元件通常编码一个兼具核酸内切酶(EN)和逆转录酶(RT)活性的蛋白质以及一个RNA分子,通过形成蛋白核酸复合物(RNP)来执行在宿主基因组中的逆转座扩增。
GII内含子可分为A、B、C三类,其中GII-C型内含子的大小仅为GII-A和GII-B的一半,被认为在进化上更古老。此前有研究发现,存在天然的缺乏开放阅读框(ORF)的GII内含子,它们被认为是GII内含子的“退化和无功能后代”,并且缺乏表征。迄今为止,广泛使用的GII内含子数据库仅列出了42个经生物信息学分析确定的缺乏ORF的GII-A和GII-B,尚未报道缺乏ORF的GII-C内含子。
刘俊杰团队对公开可用的细菌基因组序列进行初步筛选,发现许多缺乏ORF的GII-C型内含子,其中一些还在细菌基因组中存在多个拷贝,这表明,这些内含子编码的为唯一组分——RNA分子,可能在不依赖蛋白质的情况下仍然具有扩增位点识别和切割DNA的能力,从而通过“复制-粘贴”机制在基因组中进行拷贝扩增。
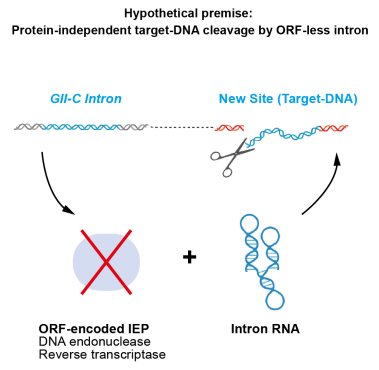
不含蛋白组分的GII内含子RNA在基因组中“复制-粘贴”的假设示意图
进一步研究显示,这些长度约600nt的RNA分子在广谱的离子浓度和温度范围内,具有显著的RNA和DNA水解切割活性。因此,研究团队将这些RNA分子命名为水解型内切核酶(Hydrolytic Endonucleolytic Ribozyme,HYER),这是科学界首次报道具有DNA水解切割能力的核酶(Ribozyme)。
研究团队利用冷冻电镜获得了HYER1的高分辨三维结构(3.0Å分辨率),并解析了其水解切割DNA的机制,HYER1通过一段暴露的单链RNA(6nt)区域——目标识别位点(TRS),来识别并招募DNA底物,将DNA捕获在结构域V所形成的催化核心中,通过经典的双镁离子机制催化DNA水解,这表明了HYER是一种TRS引导的序列特异性DNA内切酶,其主要切割位点位于TRS-DNA互补区下游0或1 nt处。
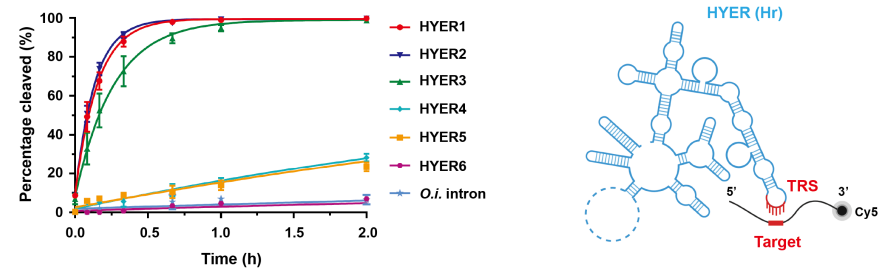
HYER的DNA切割能力
研究团队进一步验证了HYER在细胞内的DNA切割能力,在大肠杆菌和人HEK293T细胞内分别构建了ccdB毒蛋白报告系统和移码嘌呤霉素抗性基因(puro*)报告系统。结果显示,HYER可以在细胞内,尤其是真核细胞基因组中引入DNA双链断裂(DSB)并产生编辑效果。
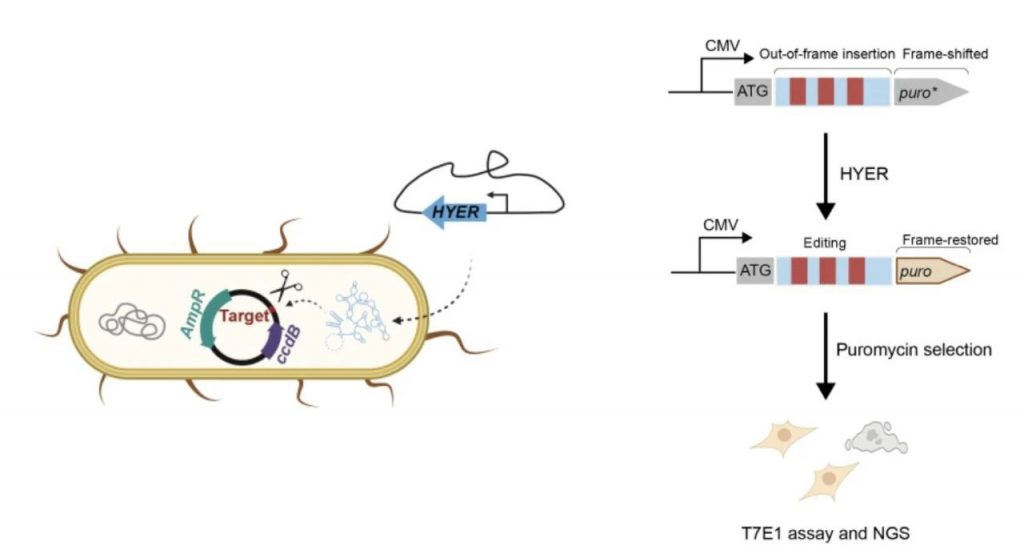
HYER在大肠杆菌和人类细胞中切割DNA双链
天然HYER仅使用TRS识别6nt的DNA区域,这限制了其精准基因编辑的能力。研究团队基于HYER的三维结构,进行了多种理性设计。例如,可根据底物序列,灵活设计目标识别位点(TRS)的序列和长度;在TRS临近区域插入14nt的底物招募序列(Recruiting Sequence,RS),可明显提高HYER1的底物识别特异性和切割效率;对回文序列和TRS进行改造, HYER1则可形成带有两个不同TRS的异源二聚体,靶向双链DNA底物的不同区域,产生了具有5’突出、3’突出或平末端的定制化切割产物。这表明了HYER具有良好的可编程性,具有相当大的改造空间。
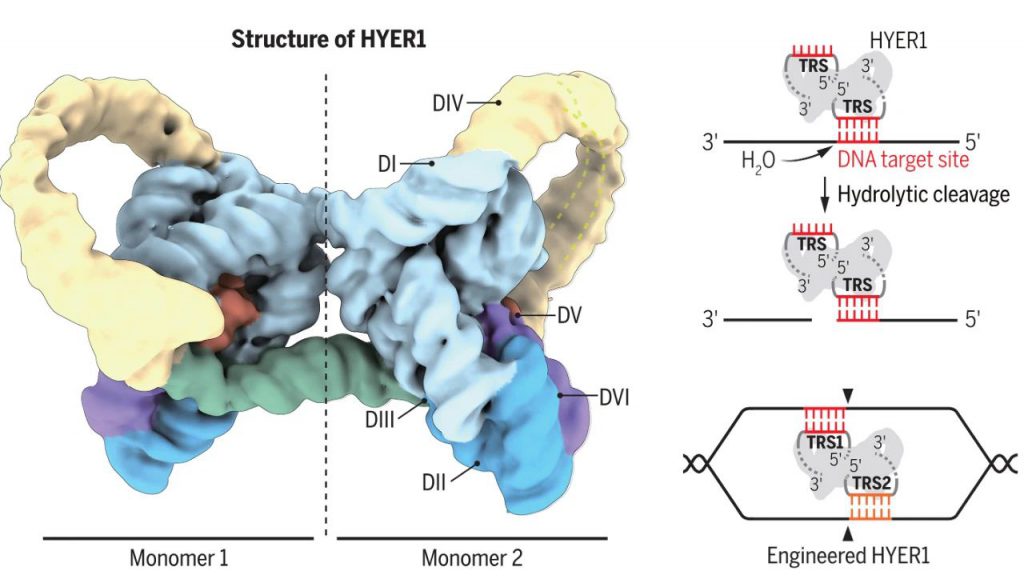
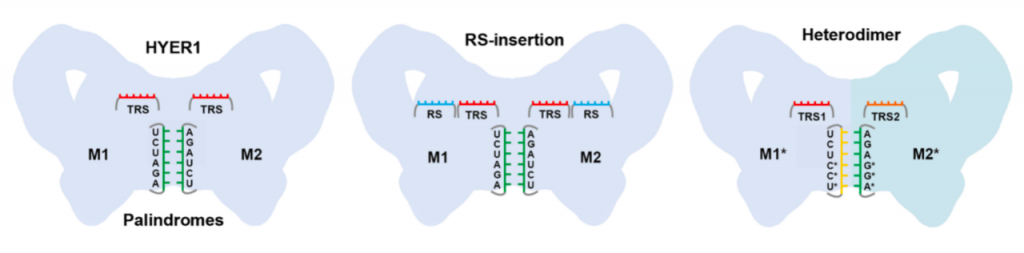
多种理性设计提高了HYER1的底物识别特异性和切割效率
最后,受“RNA世界”(RNA World)假说的启发,研究团队提出了第二类内含子(GII intron)中的“RNA的催化功能逐渐被蛋白质取代”的分子进化历程:在进化过程中,GII intron的结构域IV逐渐扩大,并产生了可编码短肽的开放阅读框(ORF),而这些短肽可作为顺式元件与内含子RNA相互作用,增强其结构稳定性和催化活性;随着ORF变得更长,更为成熟,其编码的蛋白质不仅起到稳定结构的作用,亦获得了DNA切割和逆转录活性,以替代RNA核酶行使催化功能。
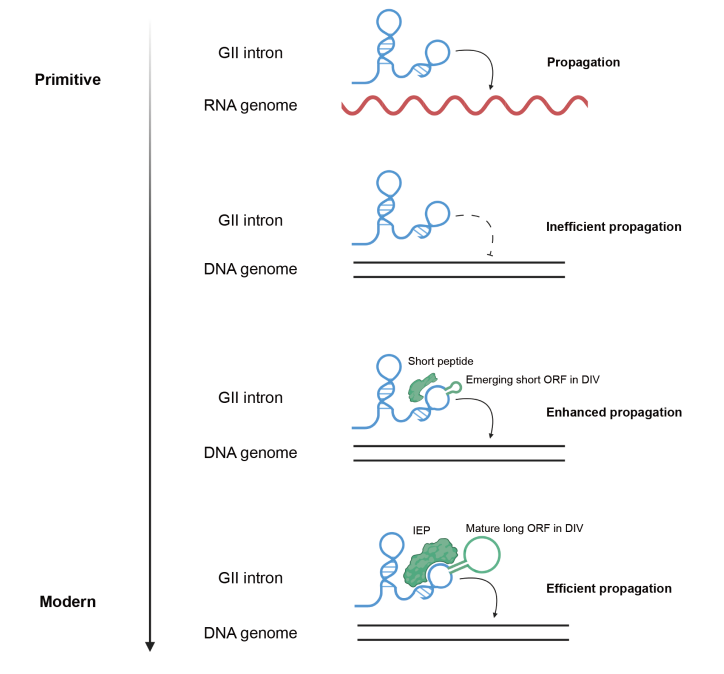
第二类内含子的进化假想
总的来说,该研究阐明了HYER的DNA靶向切割机制,其与众所周知的RNA引导的CRISPR-Cas核酸酶相似,使用可重编程的单链RNA进行DNA识别,并通过水解来切割DNA。相比广泛使用的CRISPR核酸酶的3-4kb的大小,HYER要紧凑得多,仅~0.6kb,因此更容易通过体外转录获取。
目前,天然HYER在体内编辑活性还很低,最有可能是因为其对双链DNA(dsDNA)的解旋能力有限。但正如我们在CRISPR-Cas9、CRISPR-Cas12f1等蛋白质核酸酶中所看到的那样,通过基因工程改造以提高体内基因编辑活性是可行的。
这项研究不仅拓展了科学界对RNA World假说和RNA催化功能的理解,也为创制具有中国完全自主知识产权的用于基因编辑和RNA编辑的新一代核酸操纵底盘工具奠定了基础。
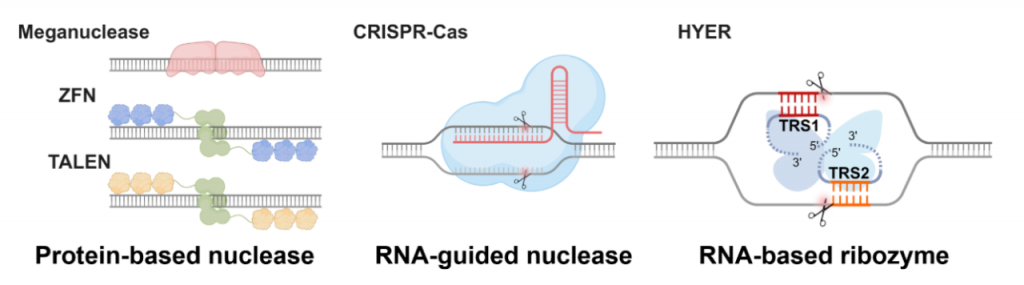
历代基因编辑工具
生物世界:您之前作为第一作者/通讯作者在Nature和Cell发表了论文,这次则是发表在了Science,投稿时有集齐CNS的考虑吗?
刘俊杰:感谢对我过往研究的关注。实际上,选择发表论文的期刊是基于多种因素的考量,每个研究项目都有其独特性。在本次的Science论文中,我认为它的主题符合Science的读者和学术范围。
生物世界:从CRISPR-Cas9以来,Base Editor以及Prime Editor都是在CRISPR的基础上做加法来实现特定的基因编辑功能,您这一次发现的HYER,可以视为一种做减法的基因编辑开发策略吗?
刘俊杰:自CRISPR-Cas9技术被开发以来,已经在基因编辑领域产生了革命性的影响。随后,基于CRISPR-Cas9的技术,如Base Editor和Prime Editor等,进一步扩展了基因编辑的可能性,它们通过对CRISPR-Cas9系统的改造,提供了更为精确和多样化的编辑功能。
在我们这项最新研究中,HYER技术的开发实际上受到了RNA World假说的启发,该假说提出,在早期生命的演化过程中,RNA可能是第一个承担遗传信息和催化反应的分子,强调RNA在生命起源和早期演化中的核心作用,尤其是具有催化生化反应的能力等。作为RNA enthusiasts,我们相信“RNA Can Do Anything”。与CRISPR-Cas系统不同,HYER摆脱了蛋白组分,其核心在于利用RNA分子本身的独特属性,如能够通过互补配对精确识别特定的DNA序列、在空间结构上具有良好的可塑性等,来实现简便的定制化核酸操纵。
通过结合对RNA功能的深入理解,HYER技术实际上是在应用RNA World假说中的原理,即利用RNA分子的多功能性和可编程性,来实现核酸操纵。这不仅表明了“RNA世界”假说在现代生物技术中的应用潜力,也进一步证实了RNA在生物体中扮演的重要角色。通过这种方式,HYER技术不仅是一种“做减法”的基因编辑策略,也是对RNA功能深入探索和应用的一个范例。
生物世界:您之前参与开发的CasX已经进行了产业转化,后续开发的这些基因编辑工具您有转化的打算或计划吗?
刘俊杰:CasX的产业转化是一个很好的例子,展示了如何将基础科学研究成果应用于实际问题解决和产业发展。对于HYER技术,我认为它在基因治疗、基因功能研究和精准农业等领域具有巨大的应用潜力。目前我们正在和多个课题组合作,进一步优化它的工作效率,拓展应用场景和潜力。
生物世界:基因编辑领域的竞争异常激烈,您的实验室率先开发出基于核酶的基因编辑工具的最重要的因素有哪些?
刘俊杰:基因编辑领域的确竞争激烈,科学家们都在努力开发各种核酸操纵工具,并且做出了很多很好的工作,比如高彩霞老师课题组不久前在 Cell 发表的基于三维结构寻找新型核酸修饰模块的工作(点此了解论文详情)【5】。我的实验室之所以关注RNA核酶并能开发它的基因编辑潜力有以下几个原因:
1)RNA World假说:如我之前所说,“RNA Can Do Anything”。我们相信自然界中存在着可以水解切割DNA的RNA分子,继而关注到不含蛋白组分,但以多拷贝形式存在的第二类内含子。我的团队成员也是RNA World假说的支持者,积极对这些第二类内含子开展体外体内活性验证,最终开发了HYER技术。
2)团队成员的多学科合作:我的团队成员来自不同的学科背景,包括生物化学、生物信息学、结构生物学和细胞生物学等。大家通力合作,多背景交叉,使得我们能够寻找并深入研究新的系统,提出并探索新颖的科学问题。
3)有力的平台支撑和经费支持:清华大学的生物计算平台和冷冻电镜平台为本研究提供了设备和技术支持,使得我们能够从宏基因组数据中大规模挖掘新的HYER系统,获得高分辨的HYER结构,继而顺利开展了一系列的优化工作。我们也非常感谢基金委原创项目(32150018)、农业部和清华大学对我们持续的经费、资源支持。
生物世界:HYER后续的改进方向有哪些?
刘俊杰:HYER技术作为一种新型的基因编辑工具底盘,虽然已经显示出了其潜力和优势,但仍有进一步改进和优化的空间。未来的改进方向至少有以下几个方面:
1)进一步提高编辑效率和特异性:虽然我们已经尝试了延长TRS、引入新的底物识别基元RS、构建异源二聚体等多种理性设计,并在成功提高了DNA切割效率和特异性,但进一步提高其编辑效率和特异性仍是重要的研究方向。这会涉及对核酶的进一步工程化改造,以优化其对目标序列的识别和切割能力。
2)融合DNA解链元件:HYER展现出了真核细胞内的基因编辑能力,但目前效率仍然较低。在富集筛选条件下,效率可达9.18%,但在不富集的条件下,效率仅为0.09%-0.2%,这表明HYER的真核基因编辑能力还有很大的进步和优化空间。我们推测这与HYER无法对双链DNA进行解旋有关,目前也在积极尝试融合DNA解链元件,以优化其真核细胞基因编辑能力。
3)提高递送效率:基因编辑技术的另一个挑战是如何高效、安全地将编辑工具递送到目标细胞或组织。评估HYER的递送效率、毒性,建立高效而安全的HYER递送方式,如使用病毒载体或LNP等,可为HYER未来的多场景应用打好基础。
4)广泛挖掘RNA核酶分子:HYER的工作表明RNA核酶的巨大应用潜力和价值,我们将进一步挖掘具有高特异性、高DNA切割活性的RNA核酶分子。
当然,我们的认知和能力是有限的,所以也非常希望科学界同行能和我们一起来拓展和探索HYER的无限可能。
清华大学生命学院刘俊杰副教授为该论文通讯作者;清华大学生命学院博士生刘子贤、博士后张寿悦、博士生朱汉舟、博士后陈之航、博士生杨韵和博士生李隆骐为该论文共同第一作者;此外,该项研究工作得到了中国科学院遗传与发育生物学研究所高彩霞课题组的大力支持。该工作由基金委原创项目(32150018)、农业部和清华大学提供经费、资源支持。
清华大学刘俊杰课题组长期关注核酸酶机制研究及核酸操纵工具的开发、应用。综合运用生物信息学、结构生物学、生物化学和细胞生物学手段,刘俊杰课题组及合作者已鉴定并开发了多种基因编辑工具(Cell, 2023;Nature, 2019;Mol. Cell, 2022;Cell Res., 2023)。课题组长期招募对新型核酸操纵系统挖掘和RNA生物学感兴趣,具有生物信息学、细胞生物学等学科背景的博士后。(实验室网页http://gogolab.life.tsinghua.edu.cn)。
论文链接:
1. https://www.science.org/doi/10.1126/science.adh4859
2. https://www.nature.com/articles/s41586-019-0908-x
3. https://www.nature.com/articles/s41422-022-00771-2
4. https://www.cell.com/cell/fulltext/S0092-8674(23)00584-6
5. https://www.cell.com/cell/fulltext/S0092-8674(23)00593-7
关于派真
作为一家专注于AAV 技术十余年,深耕基因治疗领域的CRO&CDMO,派真生物可提供从载体设计、构建到 AAV、慢病毒和 mRNA 服务的一站式解决方案。凭借深厚的技术实力、卓越的运营管理和高标准的服务交付,我们为全球客户提供一站式CMC解决方案,包括从早期概念验证、成药性评估到IIT、IND及BLA的各个阶段。
凭借我们独立知识产权的π-alphaTM 293 细胞AAV高产技术平台,我们能将AAV产量提高多至10倍,每批次产量可达1×10¹⁷vg,以满足多样化的商业化和临床项目需求。此外,我们定制化的mRNA和脂质纳米颗粒(LNP)产品及服务覆盖药物和疫苗开发的各个阶段,从研发到符合GMP的生产,提供端到端的一站式解决方案。
