

研究亮点
主要研究结果
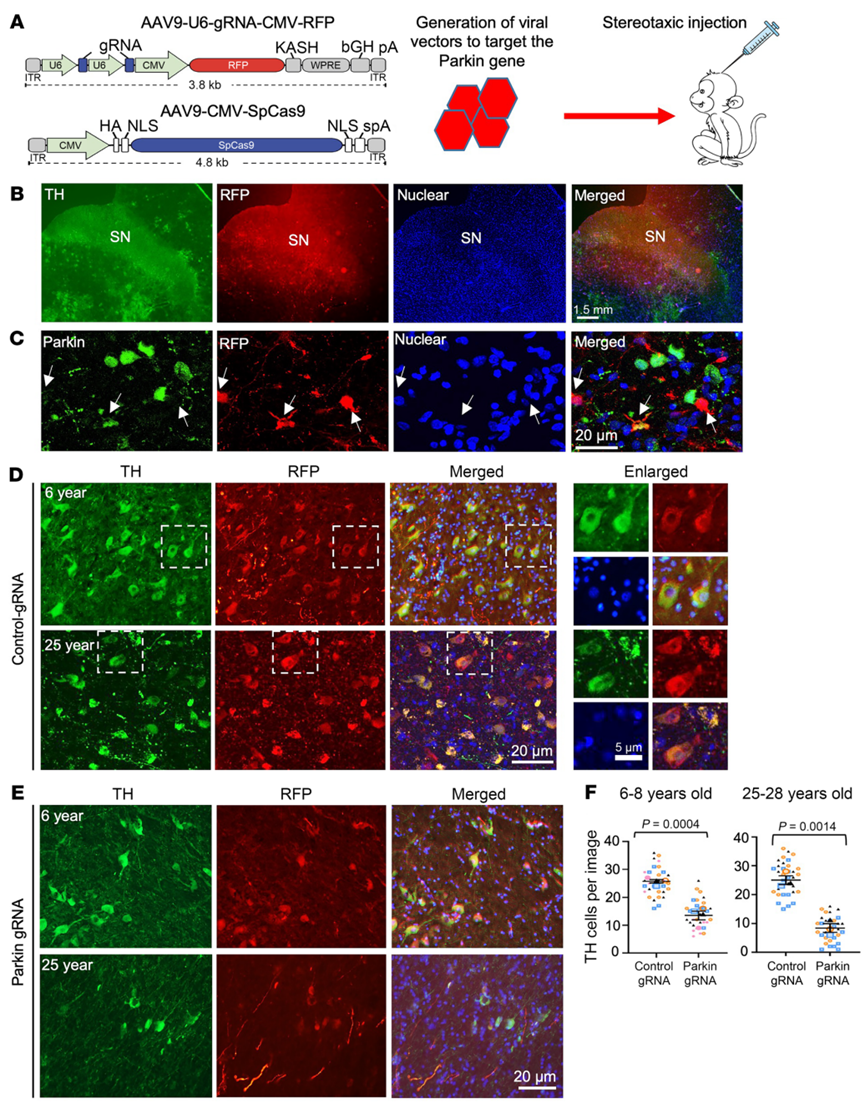
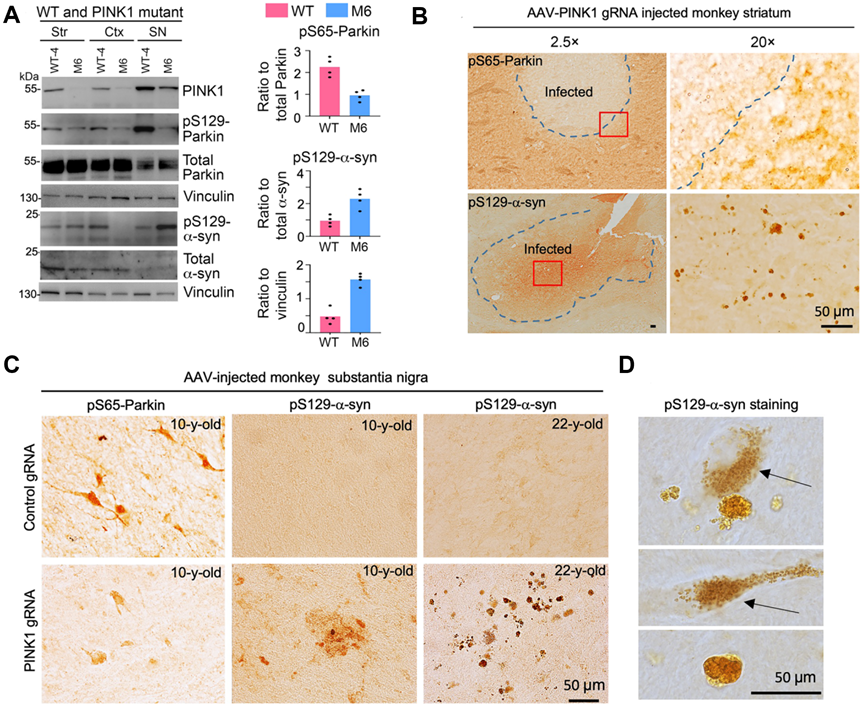
图2. PINK1缺失可导致猴中脑黑质中的Parkin磷酸化水平显著下调,病理性S129-α-synuclein蛋白表达明显增加。猴脑区注射AAV病毒载体表达CRISPR/Cas9敲除PINK1基因后,Western blot结果(A)和免疫组化染色(B-C)显示,磷酸化Parkin(pS65-Parkin)明显减少,而毒性的α-synuclein蛋白显著增加。(D)高倍镜下有毒性α-synuclein蛋白形成路易小体。
3.Parkin磷酸化的作用(图3)
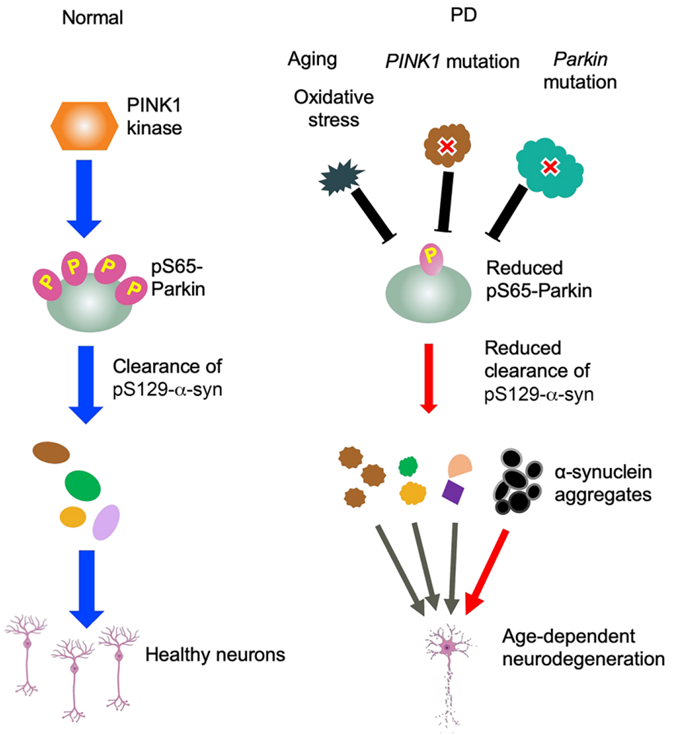
结论
| 实验动物 | 猕猴 |
| 血清型 | AAV9 |
| 启动子 | CMV, U6 |
| 注射方式 | 脑立体定位注射至前额皮质、纹状体或黑质 |
| 注射剂量 | 病毒滴度1 x 10^13 vg/mL,注射体积5~10 μL |
| 检测时间 | 感染后2-3个月 |
关于派真
作为一家专注于AAV 技术十余年,深耕基因治疗领域的CRO&CDMO,派真生物可提供从载体设计、构建到 AAV、慢病毒和 mRNA 服务的一站式解决方案。凭借深厚的技术实力、卓越的运营管理和高标准的服务交付,我们为全球客户提供一站式CMC解决方案,包括从早期概念验证、成药性评估到IIT、IND及BLA的各个阶段。
凭借我们独立知识产权的π-alphaTM 293 细胞AAV高产技术平台,我们能将AAV产量提高多至10倍,每批次产量可达1×10¹⁷vg,以满足多样化的商业化和临床项目需求。此外,我们定制化的mRNA和脂质纳米颗粒(LNP)产品及服务覆盖药物和疫苗开发的各个阶段,从研发到符合GMP的生产,提供端到端的一站式解决方案。
