慢病毒(Lentivirus)之所以被命名为“慢”,不仅源于其英文前缀“Lenti-”,更重要的是它的感染特性。这种病毒在感染初期具有较长的潜伏期且在感染后缓慢引发疾病。作为逆转录病毒科的一员,慢病毒能够感染人类和脊椎动物,主要导致巨噬细胞和淋巴细胞的原发感染,并最终引发个体发病。然而,慢病毒与其他逆转录病毒相比,其结构和基因组更为复杂,这使得慢病毒具有独特的整合机制和病毒感染持久性。
慢病毒载体的特点
慢病毒能将大量的病毒互补DNA整合到宿主细胞的DNA中,从而有效地感染非分裂期细胞,这是最有效的基因传递方式之一,更为独特的是,它们可以变成内源性的,将它们的基因组整合到宿主细胞的基因组中,使病毒的基因组可以被宿主的后代遗传。
经典的慢病毒载体(Lentivirus)是一类改造自人免疫缺陷病毒(HIV)的病毒载体,它拥有一个带有突出糖蛋白的病毒包膜,有助于附着于宿主细胞的外膜。这种病毒包含一种逆转录酶分子,它在进入细胞后对病毒遗传物质进行转录。病毒基因组内有一系列编码特定蛋白质的RNA序列,这些蛋白质有助于将病毒序列整合到宿主细胞基因组中。
与所有逆转录病毒一样,慢病毒具有gag、pol和env基因,按顺序编码病毒蛋白:5′-gag-pol-env-3’。其中,“gag”基因负责编码病毒核衣壳蛋白的结构成分,基质(MA/p17)、衣壳(CA/p24)以及核衣壳(NC/p7)蛋白;“pol”区域负责编码逆转录酶和整合酶;“env”结构域则负责编码病毒表面的糖蛋白和包膜。这些结构和编码的蛋白质在病毒感染和复制过程中起着至关重要的作用。然而,与其他逆转录病毒不同,慢病毒还具有两个调节基因:tat和rev。根据病毒的种类的不同,它们还可能具有额外的辅助基因(例如,HIV-1:vif、vpr、vpu、nef),这些基因的产物参与调节病毒RNA的合成、加工以及其他复制功能。这些额外的基因和蛋白质为慢病毒的感染和复制提供了更多的调控机制和灵活性。
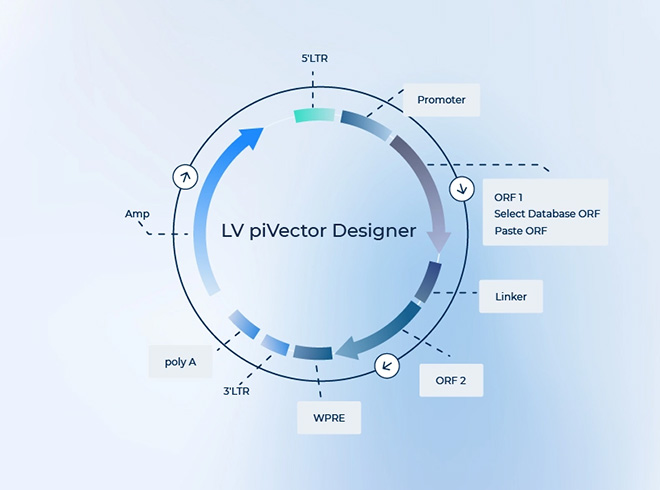
慢病毒载体在改造过程中剔除了毒性基因,保留了病毒的主要结构,使其能够高效感染和转导宿主细胞。同时,它具有长末端重复序列(LTR),可以促进病毒颗粒的包装和复制,并包含包装信号、多聚蛋白加工酶切位点和包膜蛋白等结构,以确保病毒颗粒的正确组装和释放。
经过修饰后,慢病毒载体可作为载体向细胞内插入目的基因。与其他逆转录病毒不同,慢病毒可以穿透核被膜,因此无论细胞是否正在分裂,它们都可以感染细胞,这主要是由于其衣壳蛋白的特性。许多类型的细胞(如神经元)在成体生物体中不会分裂,因此慢病毒基因疗法是治疗影响这些类型细胞的疾病的良好候选药物。
慢病毒载体的应用
正是由于慢病毒感染范围广、可实现目的基因的长时间稳定表达、操作安全性高、免疫原性低下等优点,使其在临床研究上具有广阔的应用前景。更重要的是,慢病毒载体用于改造T细胞进行CAR-T细胞治疗和基因治疗,使CAR-T细胞治疗更经济且安全有效。
关于派真
作为一家专注于AAV 技术十余年,深耕基因治疗领域的CRO&CDMO,派真生物可提供从载体设计、构建到 AAV、慢病毒和 mRNA 服务的一站式解决方案。凭借深厚的技术实力、卓越的运营管理和高标准的服务交付,我们为全球客户提供一站式CMC解决方案,包括从早期概念验证、成药性评估到IIT、IND及BLA的各个阶段。
凭借我们独立知识产权的π-alphaTM 293 细胞AAV高产技术平台,我们能将AAV产量提高多至10倍,每批次产量可达1×10¹⁷vg,以满足多样化的商业化和临床项目需求。此外,我们定制化的mRNA和脂质纳米颗粒(LNP)产品及服务覆盖药物和疫苗开发的各个阶段,从研发到符合GMP的生产,提供端到端的一站式解决方案。