
Cas12a基因敲入小鼠在多重基因组编辑、疾病建模和免疫细胞工程中的应用
研究概述
2025年3月20日,耶鲁大学助理教授陈斯迪博士在Nature Biomedical Engineering期刊发表标题为“Cas12a-knock-in mice for multiplexed genome editing, disease modelling and immune-cell engineering”的研究论文,该研究主要开发了多种Cas12a基因敲入小鼠模型,这些小鼠模型能够实现多重基因编辑、疾病建模和免疫细胞工程,并为复杂基因相互作用的解析提供了强大的工具。
研究团队在C57BL/6背景的小鼠中,通过Rosa26位点插入LbCas12a或高保真增强型AsCas12a,构建了条件性或组成性表达Cas12a的小鼠模型。这些小鼠的Cas12a组成性表达未导致可辨识的病理变化,并且能够高效实现多重基因组工程。作者团队利用这些小鼠模型进行了多种应用,包括对CD4⁺和CD8⁺ T细胞、B细胞和骨髓来源的树突状细胞进行逆转录病毒介导的免疫细胞工程;通过腺相关病毒递送多个CRISPR RNA作为单个阵列,实现内源性癌症建模;以及利用脂质纳米颗粒靶向编辑肝脏组织。此外,本研究还开发了一个同时激活和敲除双基因(DAKO)的系统。
这些Cas12a基因敲入小鼠模型以及病毒和非病毒递送载体,为体外和体内基因编辑、疾病建模、免疫细胞工程以及复杂基因相互作用的解析提供了一个多功能的工具包。

导读
正文
1. 条件性和组成性Cas12a基因敲入小鼠的生成
作者开发了LbCas12a和enAsCas12a基因敲入(KI)小鼠,以提高原代细胞中多重基因编辑的效率和简便性。作者将密码子优化的转基因克隆到Ai9 Rosa26靶向构建体中,以指导Rosa26位点第1外显子和第2外显子之间的重组(图1a)。
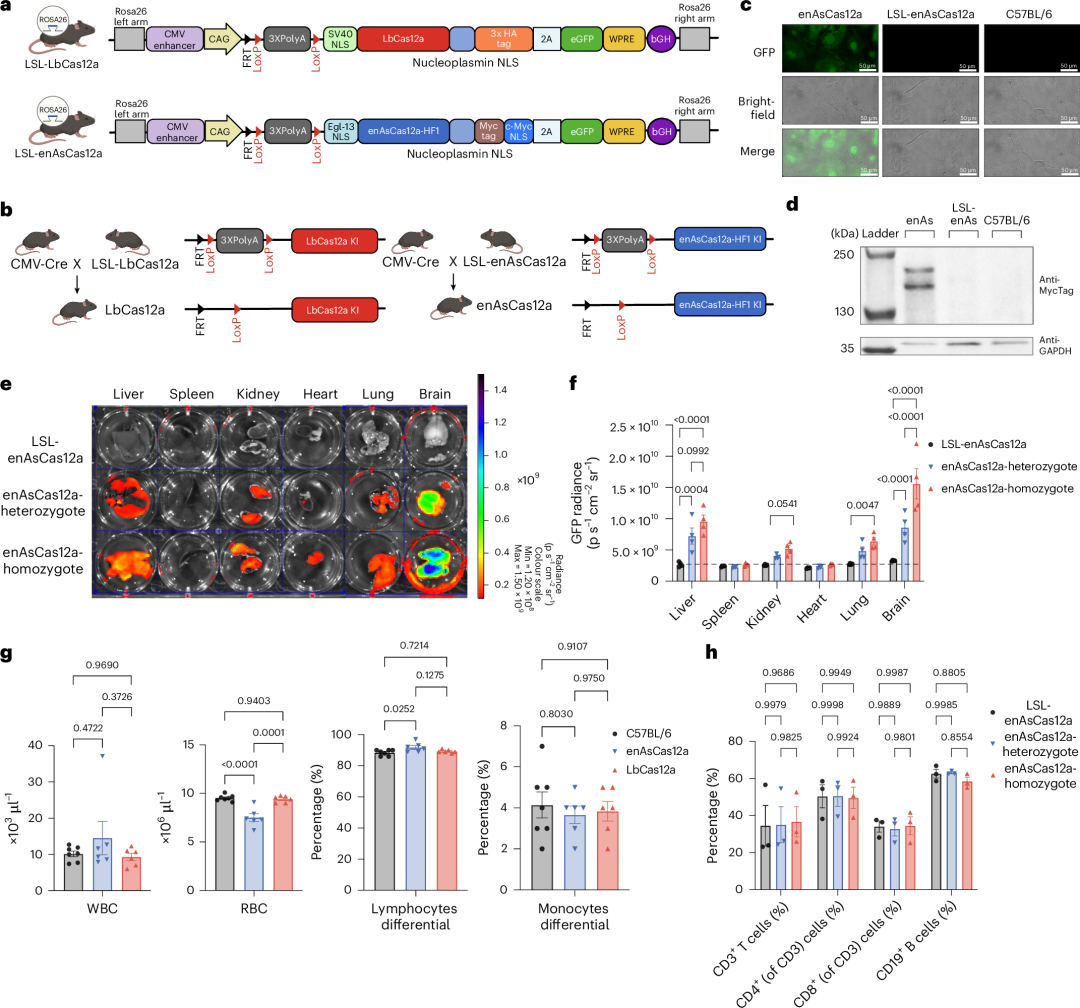
图1. 条件性和组成性LbCas12a和enAsCas12a基因敲入小鼠的生成。
2. LbCas12a小鼠实现原代免疫细胞的多重基因敲除
作者利用LbCas12a小鼠展示了其在原代免疫细胞中进行多重基因编辑的能力。由于缺乏专门针对LbCas12a的crRNA设计算法,作者通过测试不同算法设计的向导RNA,发现Broad CRISPick算法(原本为AsCas12a设计)在筛选后可显著提升LbCas12a的crRNA功能性。通过筛选CRISPick设计的crRNA,其“目标切割百分比”参数达到40%或更高,crRNA的平均切割效率提高了2.49倍,72.9%的crRNA表现出成功的基因编辑能力。这表明该算法可用于预测LbCas12a的基因编辑效率。
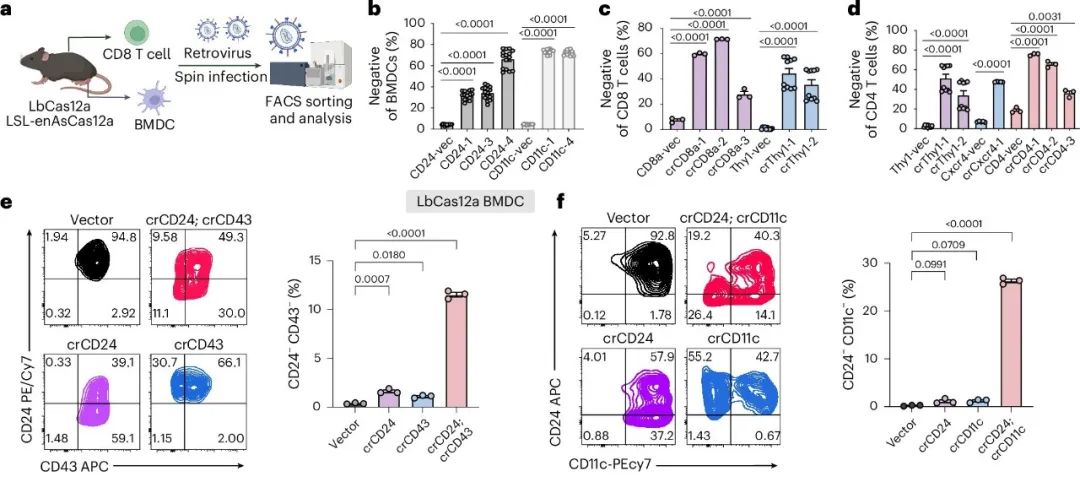
图2. 利用LbCas12a小鼠和enAsCas12a小鼠进行多重免疫细胞基因编辑。
3. LSL-enAsCas12a小鼠实现原代免疫细胞的多重体外基因工程
作者评估了LSL-enAsCas12a小鼠在原代免疫细胞中进行多重基因工程的能力。作者在BMDCs和CD8⁺ T细胞中测试了双重基因编辑,通过流式细胞术检测高表达的表面标记(图2a)。为了诱导enAsCas12a表达,作者生成了一个带有Cre和mScarlet表达盒的新逆转录病毒载体(图2g)。在BMDCs中,DKO组合crCD24-crCD11c在细胞群体水平上平均诱导了79.2%的CD24蛋白敲除和62.9%的CD11c敲除,与SKO对照相当。在单细胞水平上,crCD24-crCD11c产生了高效的双重敲除,平均有53.5%的细胞为CD24⁻;CD11c⁻(双阴性)群体(图2h)。靶向测序显示,DKO组(crCD24-crCD11c)分别诱导了CD24和CD11c基因93.8%和86.5%的基因修饰(图2i)。
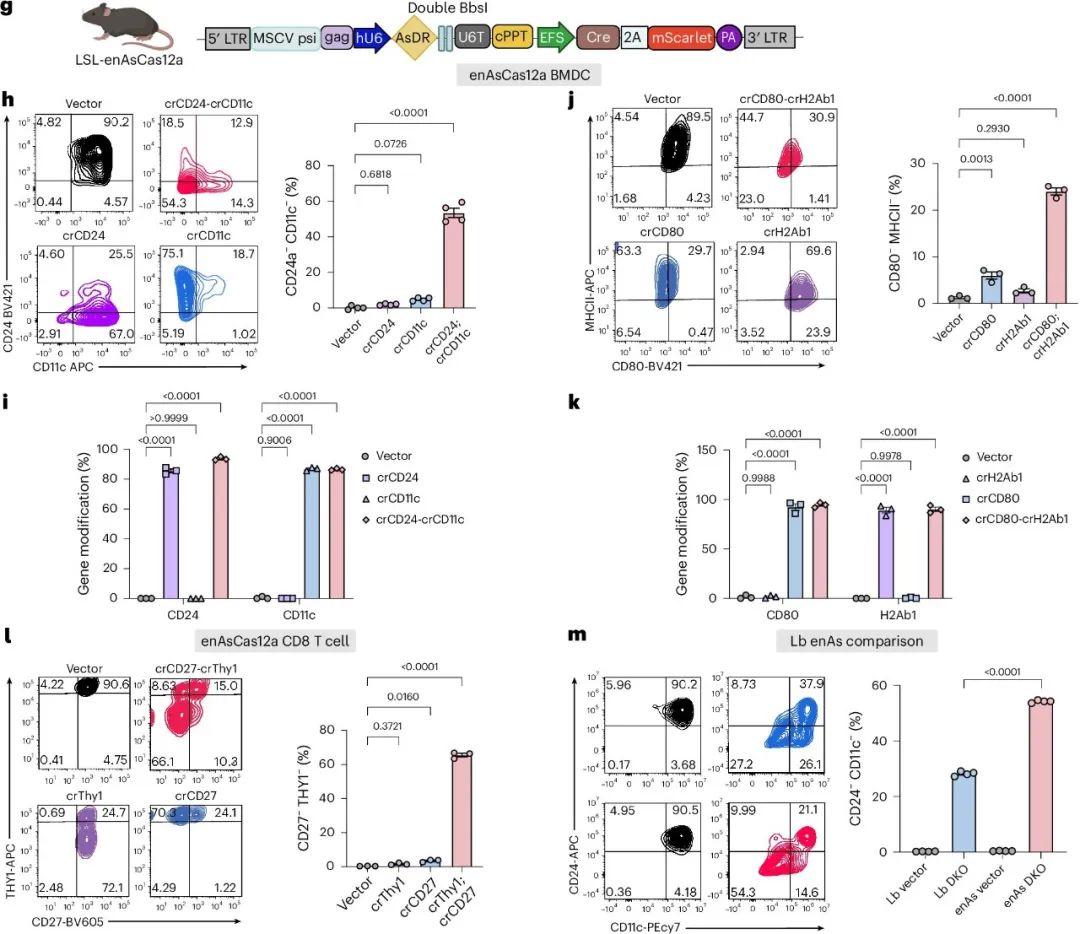
图2. 利用LbCas12a小鼠和enAsCas12a小鼠进行多重免疫细胞基因编辑。
4. 利用LSL-enAsCas12a小鼠进行高效的体内基因编辑
非病毒载体如脂质纳米颗粒(LNP)常用于体内基因编辑和治疗。作者通过静脉注射LNP-crRNA到组成性enAsCas12a小鼠体内,靶向小鼠Ttr基因(图3a),其人类同源基因是转甲状腺素蛋白淀粉样变性的治疗靶点。实验中,Ttr-cr1和Ttr-cr2两个向导RNA从注射后第6天开始显著降低血清TTR蛋白水平,且在20天监测期内保持稳定(图3b)。Ttr-cr1几乎实现了100%的血清TTR水平敲除,Ttr-cr2也达到了近50%的敲除效果。
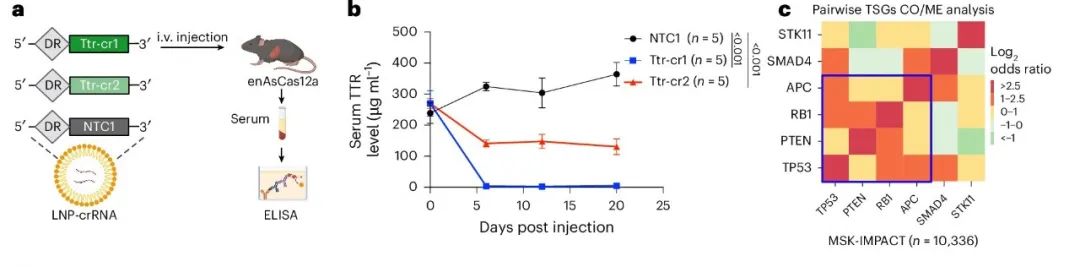
图3. 通过肝脏LNP-crRNA靶向和AAV介导的肿瘤发生展示的体内基因编辑。
5. LSL-enAsCas12a小鼠实现体内多重基因编辑用于自发性癌症建模
自发性癌症模型通过在完整生物体内从正常细胞直接诱导肿瘤,更接近人类肿瘤发生过程和微环境。传统方法构建基因工程小鼠模型过程繁琐,而CRISPR基因组工程,尤其是Cas9基因敲入小鼠及其病毒递送方法,简化了自发性癌症建模。然而,Cas9系统在肿瘤建模中因缺乏多重基因编辑能力和AAV载体包装尺寸限制而受限。Cas12a具有高多重化能力,可通过单个载体中的crRNA阵列编码多个crRNA靶向多个基因。因此,LSL-enAsCas12a小鼠成为更适应性强和精确的自发性癌症建模的高效工具。
作者从泛癌数据库中选择了六个经常发生突变的肿瘤抑制基因(TSGs),经过分析后选择了TP53、APC、PTEN和RB1。为了同时靶向小鼠的Trp53、Pten、Apc和Rb1,作者克隆了一个串联的RNA阵列(crTSG),由单个U6启动子驱动(图3d)。Cre重组酶在EFS启动子下表达,以激活enAsCas12a-HF1-eGFP表达。通过静脉注射AAV-crTSG-Cre,100%的小鼠在一个月内发展出侵袭性头颈部癌症,而对照组没有癌变(图3e)。肿瘤切片在免疫组化分析中显示出enAsCas12a-HF1-GFP表达,表明肿瘤是由AAV-crTSG-Cre介导的基因编辑诱导的(图3f)。
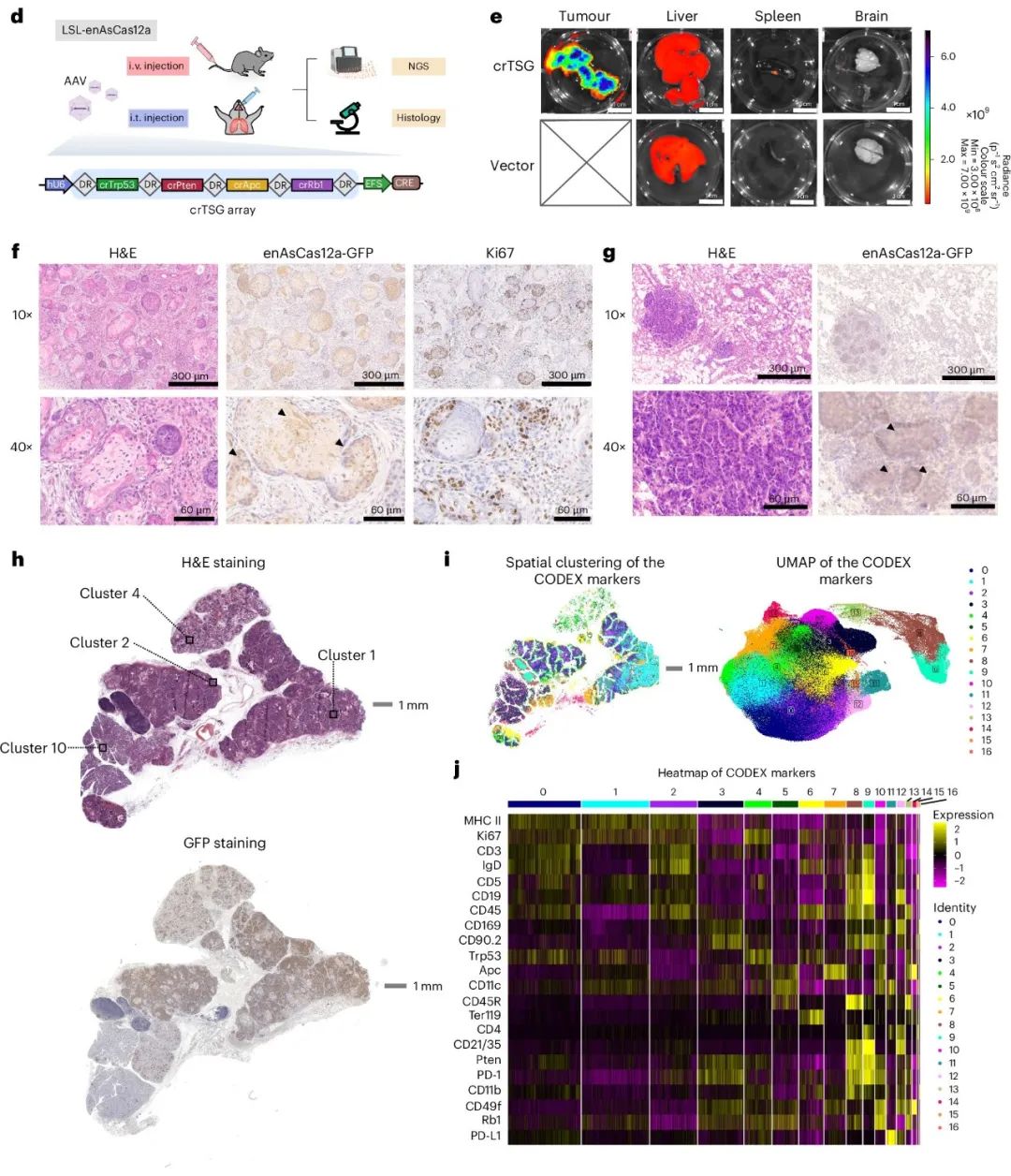
图3. 通过肝脏LNP-crRNA靶向和AAV介导的肿瘤发生展示的体内基因编辑。
作者进一步使用CODEX技术对SCC进行了表征,该技术能够在单细胞分辨率下进行多重蛋白质组学分析(图3h-j)。基于22个标记物的表达水平,SCC细胞被分解为17个簇。这些肿瘤区域表现出高度的异质性,与通过癌细胞系建立的异种移植肿瘤相比,这种模型在生理学上更接近人类肿瘤。总之,LSL-enAsCas12a小鼠展示了体内多重基因组工程的稳健性,使得自发性癌症建模成为可能。
6. 肿瘤和免疫细胞DKO样本的基因组分析
作者通过基因组水平的分析,量化了LSL-enAsCas12a小鼠的on-target和off-target效率。对鳞状细胞癌(SCC)样本进行的NGS显示,4个靶向肿瘤抑制基因(Trp53、Apc、Pten和Rb1)的基因修饰水平显著(平均约50%),且大多数修饰为缺失,集中在预测的enAsCas12a crRNA切割位点附近(图4a、b)。在单细胞水平上,10个肿瘤单克隆中有10个在两种癌症类型中都对所有四个基因进行了修饰(图4c、d)。
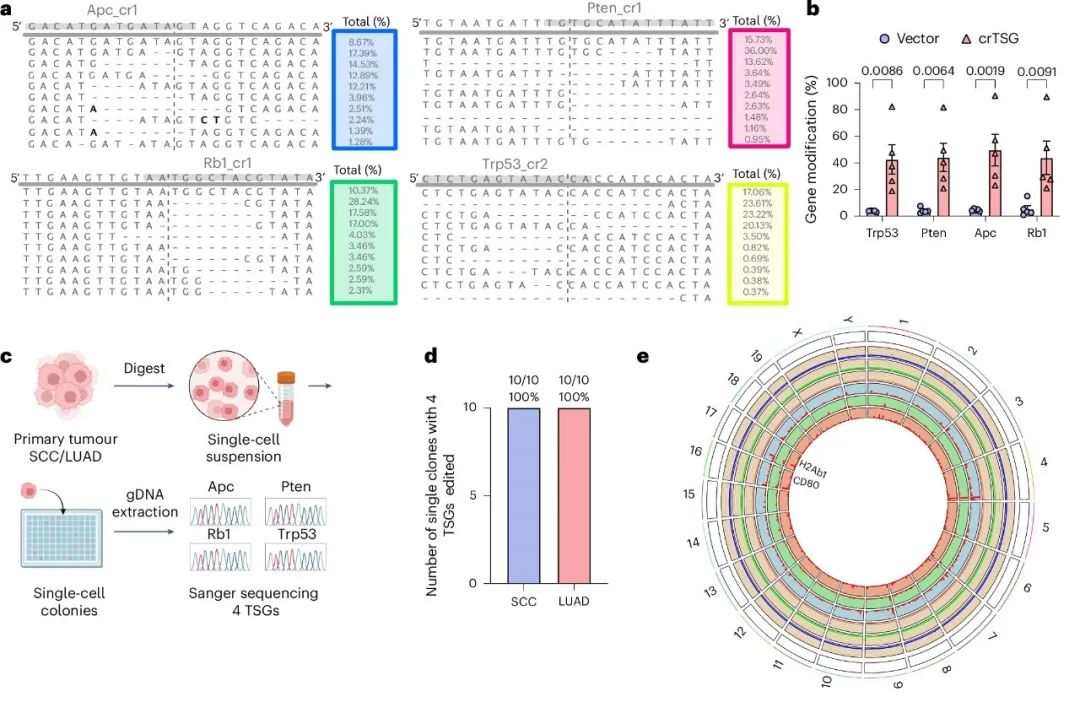
图4. enAsCas12a基因敲入小鼠显示出高靶向性和低脱靶性的编辑效率。
作者通过全基因组测序(WGS)和靶向扩增子测序来表征LSL-enAsCas12a小鼠在CD80-H2Ab1 DKO实验中的off-target率。WGS显示,DKO和SKO的独特变异在多个染色体上存在变异热点(图4e)。尽管WGS能够对整个基因组中的潜在off-target位点进行调查,但其测序深度有限。因此,作者采用靶向扩增子测序作为替代方法,其读取深度要高得多。结果显示,少数预测的off-target位点(crCD80_4、crCD80_10和crH2Ab1_10)上观察到修饰,但这些位点在载体组中也发现了修饰,表明这些indels可能由PCR偏差引起(图4f-j)。
这些结果表明,LSL-enAsCas12a在保持可控的off-target效应水平的同时,实现了高效的on-target多重基因编辑。
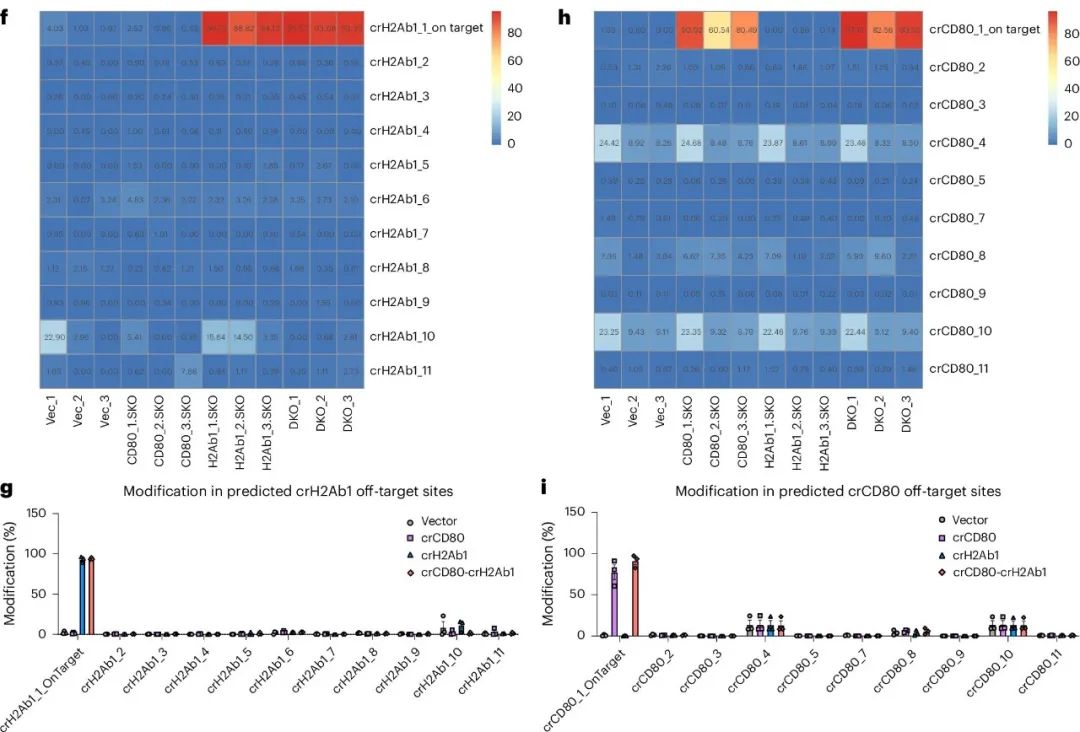
图4. enAsCas12a基因敲入小鼠显示出高靶向性和低脱靶性的编辑效率。
7. 利用LSL-enAsCas12a-HF1小鼠开发同时激活和敲除基因(DAKO)系统
为了在单个细胞中同时实现基因激活和敲除,作者开发了DAKO系统。通过将LSL-enAsCas12a-HF1小鼠与dCas9-SPH小鼠杂交,生成了LSL-enAsCas12a-HF1;dCas9-SPH DAKO小鼠(图5a)。
作者设计了一种Cas9–Cas12a融合向导RNA系统,该系统在单个序列中表达两种类型的CRISPR向导RNA(Cas9-sgRNA和Cas12a-crRNA),通过AsCas12a DR序列连接,用于在相同细胞中同时表达(图5b)。这种向导RNA嵌合体被enAsCas12a-HF1蛋白切割,生成成熟的Cas9-sgRNA和Cas12a-crRNA,分别指导dCas9-SPH进行基因激活和enAsCas12a-HF1进行基因敲除。
为了验证该系统,作者从DAKO小鼠中分离出BMDCs,并用携带DAKO融合向导盒(包含Itgb4-sgRNA与AsDR-crCD24连接)的逆转录病毒以及组成性活性EFS-Cre盒(sgItgb4-crCD24-Cre载体)感染它们(图5c)。

图5. 利用LSL-enAsCas12a-HF1;dCas9-SPH双转基因小鼠实现的同时激活和敲除(DAKO)。
流式细胞术分析显示,sgItgb4-crCD24-Cre DAKO靶向在细胞群体水平上导致CD24蛋白减少,与单敲除对照相当(图5d、e)。同时,DAKO组中ITGB4蛋白在细胞群体水平上上调,表明基因激活成功。DAKO组中观察到预期的双重靶向细胞群体,其基因调控方向正确(图5d、e)。这些结果表明,DAKO系统能够在单细胞水平上实现高效且同时的基因激活和敲除,为免疫细胞中的正交双基因靶向提供了新的工具。
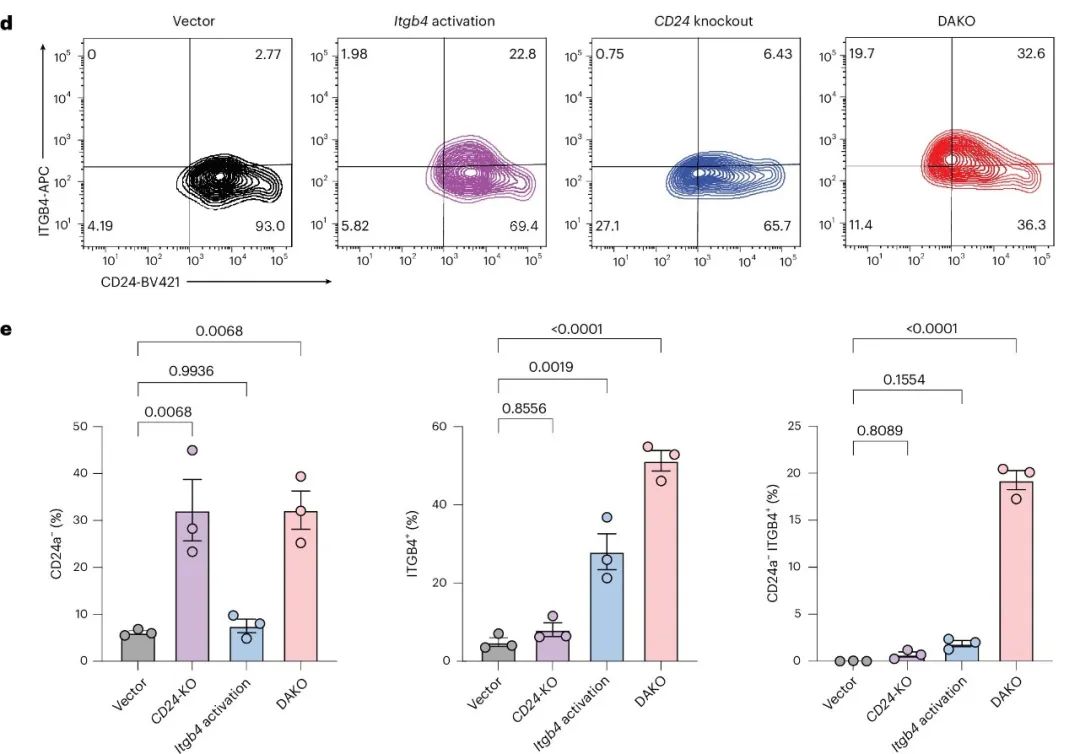
图5. 利用LSL-enAsCas12a-HF1;dCas9-SPH双转基因小鼠实现的同时激活和敲除(DAKO)。
展望
解析复杂基因调控网络对于理解基础生物学和人类疾病的多效性至关重要。因此,开发一种多功能的体内和体外多重基因组工程技术是系统研究基因相互作用的必要条件。本研究成功培育了四种多功能小鼠品系:条件性LSL-LbCas12a和组成性LbCas12a小鼠,以及条件性LSL-enAsCas12a和组成性enAsCas12a小鼠。LbCas12a和enAsCas12a基因敲入小鼠及其配套的递送系统,可作为一套多功能的新型遗传工具。将多个crRNA整合到单个crRNA阵列中进行多重基因扰动,简化了克隆过程。作者发现,LbCas12a和enAsCas12a小鼠能够在多种免疫细胞类型中实现高达90-100%的高效基因编辑,并且在单细胞水平上展示了Cas12a小鼠的多重基因编辑能力。
为了兼容条件性和组成性enAsCas12a品系,作者开发并应用了多种递送方法(脂质纳米颗粒LNP、腺相关病毒AAV和逆转录病毒),分别用于高效的肝脏基因靶向、肿瘤建模和原代免疫细胞编辑。利用LNP-crRNA在一周内高效敲除了肝脏中的Ttr基因。通过同时敲除四个肿瘤抑制基因,迅速诱导了唾液腺鳞状细胞癌(SCC)。在原代BMDCs和CD8 T细胞中展示了高效的双重敲除(DKO),其效率高于LbCas12a小鼠。作者还通过与dCas9-SPH系统整合,开发了同时激活和敲除基因(DAKO)系统,展示了转基因小鼠的模块化能力。
本研究的组成性和条件性Cas12a小鼠品系能够高效地进行体内和体外多重基因组工程,为多种领域提供了应用机会。尽管由2A肽诱导的“自切割”效率还有待提高,但这些小鼠表现出与Cas9基因敲入小鼠相当的编辑效率,并且与Cas9小鼠相比,具有基于crRNA阵列的多重基因编辑的独特优势。这些小鼠可以促进体内治疗性基因靶向、疾病/肿瘤建模和原代免疫细胞工程的快速无缝工作流程。
结合寡核苷酸文库合成和克隆,这些小鼠可用于高通量CRISPR筛选,以研究复杂的遗传问题,例如免疫细胞中效应细胞和记忆细胞表型的转变。此外,这些小鼠允许构建多种肿瘤类型(如SCC和LUAD)的自发性肿瘤模型,或具有复杂基因型的肿瘤模型(例如,不同组合的肿瘤抑制基因的插入/缺失和染色体非整倍性)。作者注意到,在组成性Cas12a品系中,不同器官中Cas12a的表达水平存在差异。因此,在使用这些小鼠进行体内CRISPR筛选时,应考虑靶组织中Cas12a的表达水平。建议使用具有组织/细胞类型特异性的Cre驱动因子来驱动Cas12a的表达。开发其他变体的Cas12a小鼠,包括此处的两种Cas12a物种,以及敲入到不同安全港位点(如H11-enAsCas12a小鼠)的Cas12a小鼠,代表了为多样化应用领域开发广泛启用的Cas12a基因编辑工具包的汇聚性集体发展。
关于派真
作为一家专注于AAV 技术十余年,深耕基因治疗领域的CRO&CDMO,派真生物可提供从载体设计、构建到 AAV、慢病毒和 mRNA 服务的一站式解决方案。凭借深厚的技术实力、卓越的运营管理和高标准的服务交付,我们为全球客户提供一站式CMC解决方案,包括从早期概念验证、成药性评估到IIT、IND及BLA的各个阶段。
凭借我们独立知识产权的π-alphaTM 293 细胞AAV高产技术平台,我们能将AAV产量提高多至10倍,每批次产量可达1×10¹⁷vg,以满足多样化的商业化和临床项目需求。此外,我们定制化的mRNA和脂质纳米颗粒(LNP)产品及服务覆盖药物和疫苗开发的各个阶段,从研发到符合GMP的生产,提供端到端的一站式解决方案。
